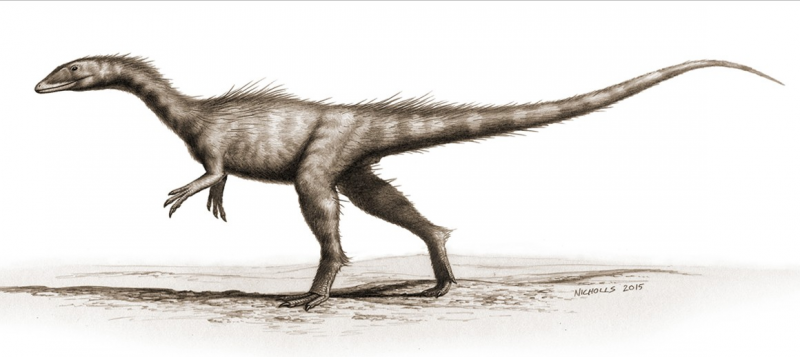
Dracoraptor hanigani (teropod) restaurerad som ett rovdjur som lever vid kusten och som asätare. Konstverk av Bob Nicholls (paleocreations.com). Kredit:Naturhistoriska museet, London. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Utvecklingen av endotermi (termoreglering med metabola medel) representerar en stor övergång i ryggradsdjurens historia. Dock, processen för endotermiutveckling och dess tidslinje hos fåglar och däggdjur är fortfarande kontroversiell. I en ny rapport om Vetenskapens framsteg , Enrico L. Rezende och ett team av forskare vid Center of Applied Ecology and Sustainability, och Institute of Environmental and Evolutionary Sciences i Chile, kombinerade en värmeöverföringsmodell med kroppsstorleksdata för theropod. Forskarna rekonstruerade sedan utvecklingen av ämnesomsättningshastigheter längs fågelstammen. Resultaten antydde att en minskning i storlek utgjorde vägen för minsta motstånd för endotermi att utvecklas - maximera termisk nischexpansion, samtidigt som kostnaderna för förhöjda energikrav minskar.
Som ett resultat, forskarna antar att metabolismen skulle ha ökat med miniatyrisering under tidig-mellanjuraperioden (för ungefär 180 till 170 miljoner år sedan) för att resultera i en gradient av metaboliska nivåer i theropod-fylogenin. Medan basala teropoder kan ha uppvisat lägre metabolismhastigheter, de nyare icke-fågelhärkomsterna var sannolikt anständiga termoregulatorer med förbättrad metabolism. Analysen gav en preliminär sekvens i tiden av de viktigaste evolutionära övergångarna, för uppkomsten av små, endotermiska och flygande fjäderdinosaurier.
Utvecklingen av endotermi hos fåglar och däggdjur är en viktig övergång under ryggradsdjurens evolution och ger ett extraordinärt exempel på evolutionär konvergens mellan grupper, avgörande för deras utbredda geografiska spridning och ekologiska framgång. Även om flera grupper av ryggradslösa djur och ryggradsdjur kan höja sina temperaturer över omgivningen, förmågan att upprätthålla en hög och konstant kroppstemperatur via endogen värmeproduktion i vila är exklusiv för fåglar och däggdjur. Observationen förklarade deras större rörlighet, uthållighet och tolerans under en rad förhållanden. Dock, denna strategi är energiskt kostsam och har inga virtuella spår i fossilregistret, därför förblir takten och sättet för endotermi kontroversiellt under ryggradsdjurens evolution.
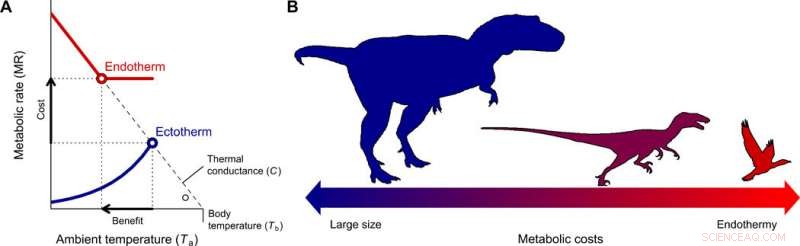
Utvecklingen av endotermi och miniatyrisering i theropodlinjen som leder till fåglar. (A) Kostnads-nyttan att byta från ektotermi till endotermi för olika kroppsstorlekar kvantifierades med Scholander-Irving-modellen, som beskriver hur en ökning av ämnesomsättningen i vila (kostnad) ökar den termiska nischen Tb − Ta (nytta). Eftersom det inte finns någon termisk gradient mellan organismen och miljön i frånvaro av värmeproduktion, denna kurva skär abskissan vid Tb =Ta när MR =0 (8). De heldragna blå och röda linjerna visar de metaboliska kurvorna för en typisk ektoterm och endoterm, respektive, och de öppna symbolerna visar den maximala termiska gradienten Tb − Ta möjlig med vilande metaboliska hastigheter, används i vår modell (Ekv. 2). (B) En minskning av kroppsstorlek, överensstämmer med den som beskrivs från förfäders theropoder till basalfåglar (22), utgör den evolutionära vägen för minsta motstånd eftersom energikostnaderna för att vara stor växlas mot de för att vara endotermiska. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
För att förstå ursprunget och orsaken till endotermi under utvecklingen av fåglar och däggdjur, forskarna övervägde två grundläggande frågor. Vilka är kostnaderna och fördelarna med denna strategi jämfört med ektotermi? Och vilka förhållanden gynnade en övergång till endotermi? Rezende et al. tog upp frågorna med hjälp av Scholander-Irvings modell för värmeöverföring, används för att studera termoreglering i endotermer i mer än 60 år. Relationen användes sällan för ektotermer, men eftersom alla levande organismer producerade endogen värme, modellen förblev tillämpbar under termiskt stabilt tillstånd. De gjorde detta avgörande antagande för att undvika att använda komplexa dynamiska modeller som ofta tillämpas på ektotermi, vilket skulle kunna försvåra föreliggande analyser.
De kvantifierade kostnaderna för endotermi som massoberoende energiutgifter, där fördelarna inkluderade ökad rörlighet och födosökseffektivitet, undvikande av rovdjur, tolerans mot och kolonisering av ett brett spektrum av miljöförhållanden, med ökade tillväxthastigheter och homeostas. Rezende et al. kvantifierade den termiska nisch som organismer kunde ockupera och dess expansion för att uppskatta nettovinsten av endotermi. De beräknade sedan kostnads-nyttan av att anta en endoterm livsstil med hänvisning till den ektotermiska förfadern och endotermiska ättlingen. Som ursprungligen föreslogs av evolutionsbiologen Bran K. McNab, när forskarna replikerade dessa beräkningar med exakta kroppsstorleksberäkningar, resultaten visade att mindre storlekar minskade energikostnaderna för att utvecklas mot endotermi.
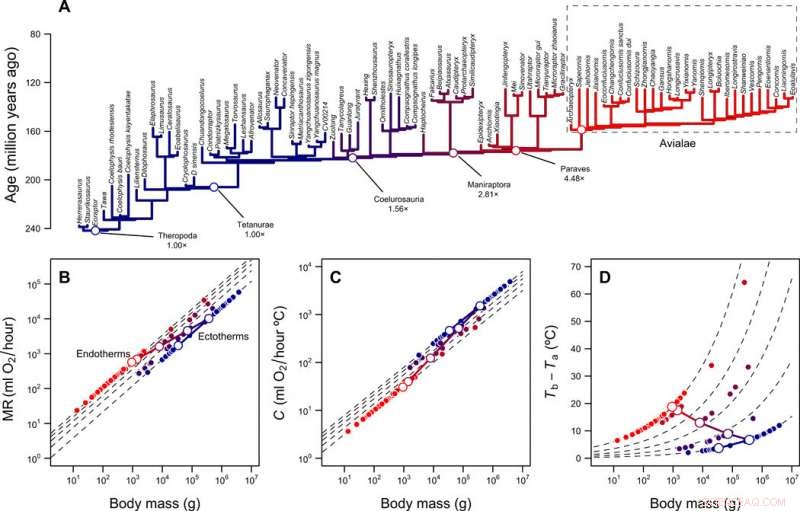
Rekonstruktion av metaboliska nivåer och termisk nisch av theropoder. (A) Theropod fylogeni med grenar färgkodade enligt rekonstruerade metaboliska nivåer. (B) Skalning av metabolisk hastighet kontra kroppsmassa för ektotermer (MR =0,68 massa 0,75) och endotermer (MR =3,4 massa 0,75) och den förutspådda banan för fågelstamlinjen under övergången från ektotermi till endotermi. Streckade linjer visar veckskillnader mellan ektotermer och endotermer (1× till 5×); öppna och stängda symboler avbildar rekonstruerade värden för fågelstammens härstamning och spetsarna av fylogenin, respektive. (C) Skalning av termisk konduktans C och kroppsmassa för ektotermer (C =2,5 mass0,5) och endotermer (C =1,0 mass0,5), veckskillnader från 2,5× till 1×. (D) Termisk gradient och veckskillnader beräknade med ekv. 1 (i publikationen) och värden i (B) och (C). Log-log linjära banor som förbinder MR och C för den ektotermiska förfadern och den endotermiska avkomman, såväl som den resulterande banan i termisk gradient, visas med genomgående linjer. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Teamet utforskade hur denna värmeöverföringsmodell kombineras med fylogenier och rekonstruktioner av kroppsstorlek, kasta ljus över utvecklingen av endotermi hos fåglar och deras förfäder för theropod. Rezende et al. uppskattade kostnaderna för att utveckla endotermi längs fågelstamlinjen med hjälp av rekonstruerade förfäders kroppsstorlekar, baserat på fossilregistret. För att kvantifiera energikostnaderna inom alternativa scenarier simulerade de utvecklingen av kroppsstorlek längs linjen och erhöll fördelningen av kostnad per grad i modellen. De antog en oriktad Ornstein-Uhlenbeck (OU) modell mellan en massa på 10 g och 100, 000 kg med en genomsnittlig utvecklingshastighet motsvarande den som rapporterats för theropoder. Simuleringarna indikerade en markant minskning av energikostnaderna per grad med miniatyrisering. Forskarna förklarade de minskade kostnaderna med hjälp av två fenomen.
Först, expansionen i termisk nisch baserad på en ökning av metabolisk hastighet (MR) var oproportionerligt högre i större ektotermer på grund av deras förmåga att upprätthålla en hög kroppstemperatur (T b ). Detta åtföljdes av en relativt låg massoberoende ämnesomsättning, på grund av initial homeothermi (stabil termoreglering). Följaktligen, de noterade att större startstorleken på den ektotermiska förfadern, billigare övergången till endotermi. Andra, under miniatyrisering, djuren bytte ut energikostnaderna för att vara stora mot att vara endotermiska. Resultaten förklarade hur de höga energiomsättningshastigheterna utvecklades, oavsett deras inverkan på mat- och vattenbehov. Trots inneboende variation i resurstillgänglighet under evolutionstid - mindre storlekar och högre energiomsättningshastigheter gynnades systematiskt i denna härstamning. Resultaten av arbetet överensstämde kvalitativt med andra modeller om utvecklingen av endotermi.
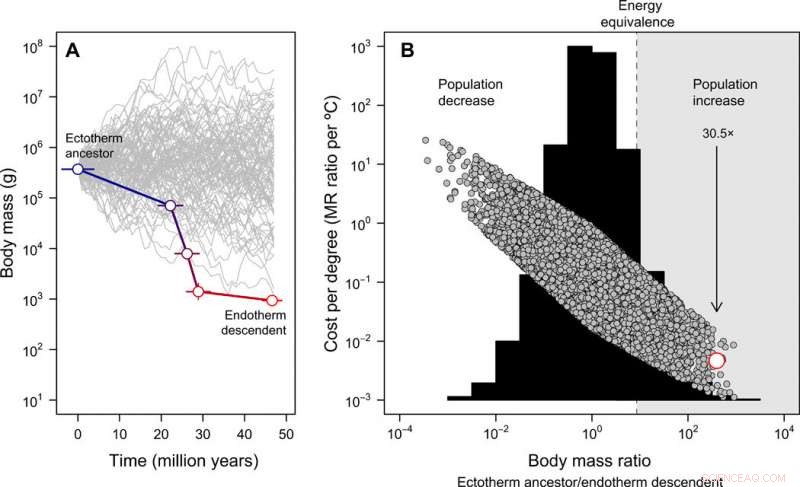
Utveckling av kroppsstorlek och kostnadsfördelarna med endotermi. (A) Miniatyriseringen från Tetanurae till basala fåglar som härleds från fossilregistret, kontrasteras mot 100 simulerade storleksbanor utgående från samma förfäders kroppsstorlek för illustrativa ändamål (observera att för den efterföljande fullständiga nollmodellen, släktkroppens storlek tillåts variera). Felet representerar SD i rekonstruerade värden över 20 kandidatträd. (B) Frekvensfördelningen av kroppsviktsförhållanden erhållna över 10, 000 simulerade kroppsstorleksbanor (histogram) och energikostnaderna för att utveckla endotermi uttryckt per grad Celsius (Ekv. 2 i publikationen) under denna nollmodell (grå symboler). I detta fall, den förfäders kroppsstorlek erhölls från en enhetlig fördelning som sträckte sig mellan 10 g och 100, 000 kg. Den empiriska uppskattningen i fågelstamlinjen visas i rött. Den region där en minskning av kroppsstorleken skulle kompensera för energikostnaderna för utvecklande endotermi, göra det möjligt för befolkningen att öka i ett scenario med konstanta resurser, är markerad i grått. Pilen visar den förväntade befolkningsökningen, med tanke på den observerade kroppsstorleksminskningen i fågelstammens härstamning när endotermi utvecklades. Dessa analyser indikerar att energikostnaderna för att utveckla endotermi minskar med miniatyrisering och, som ett resultat, befolkningsstorleken kan ha ökat trots de metabola kostnaderna för en endoterm livsstil. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Storleksminskningen i fågelstammens härstamning, nära den teoretiska vägen för minsta motstånd för att endotermi ska utvecklas, sedan Rezende et al. rekonstruerade hur fenomenet kan ha utvecklats i therapod-fylogenin. Processen indikerade ökningen av MR (metabolisk hastighet) som sträcker sig över större delen av tidig-mellanjuran (för ungefär 180 till 170 miljoner år sedan), involverade terapeutgrupper där förekomsten av protofjädrar och fjädrar redan var allestädes närvarande. De föreslog också mycket olika metaboliska hastigheter över de samtidiga linjerna av Coelurosauria, Maniraptora och Paraves, vilket skulle kunna förklara uppkomsten och diversifieringen av dessa grupper under den sena juraperioden. Arbetet kan också förklara den onormalt höga mångfalden av Coelurosauria vid mellanliggande kroppsstorlekar (30 till 300 kg) jämfört med andra dinosauriegrupper. Forskargruppen demonstrerade vidare en nischfyllande modell av adaptiv strålning i mesozoiska dinosaurier med exceptionell minskning av kroppsstorleken i fågelstammens härstamning (dvs inom basala noder av Coelurosauria och Paraves), dock, de föreslog inte en lämplig evolutionär hypotes för att redogöra för resultatet.
På det här sättet, Enrico L. Rezende och kollegor observerade två exceptionella fenomen under fåglarnas evolution; (1) en ihållande (men inte nödvändigtvis gradvis) miniatyrisering som spänner över miljoner år och (2) uppkomsten av endotermi. De använde rekonstruktioner för att föreslå åtföljande utveckling av endotermi med en minskning i storlek längs fågelstamlinjen, som också tidigare föreslagits för däggdjur. Motsvarande terapeutiska kladdar uppvisade ett helt spektrum av MR. Tolkningarna indikerar att endotermi föregick utvecklingen av flygningen och den markanta minskningen av kroppsstorleken under utvecklingen av fågelstammens härstamning åtföljdes av en stor förändring i metabola nivåer.
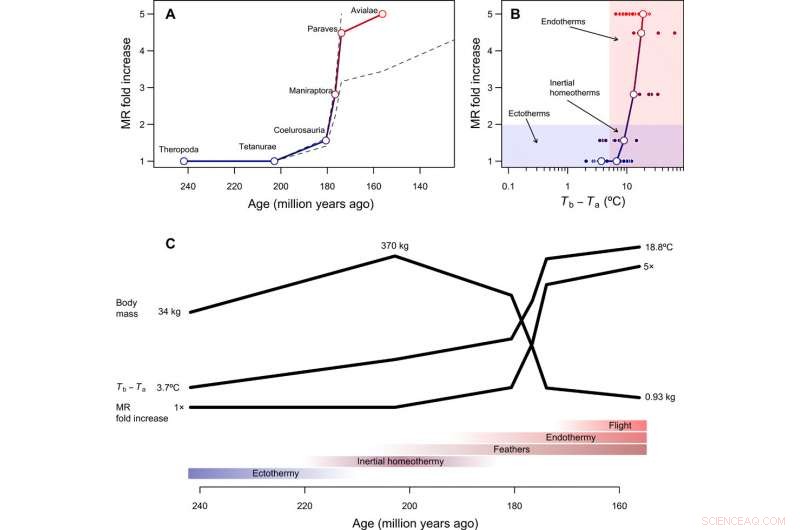
Tempo och läge i utvecklingen av endotermi. (A) Rekonstruerat temporalt förlopp av metabolisk evolution i fågelstamlinjen, med streckade linjer som visar hur rekonstruktioner förändras om man antar att antingen Paraves eller Neornithes var helt endotermiska istället för basalfågeln [för beräkningar med Neornithes, vi antog en kroppsstorlek på 150 g baserat på uppskattningar för Vegavis och en tidsuppskattning på 100 Ma sedan]. Den dubbla ökningen av MR beräknades genom att dividera den rekonstruerade MR under övergången till endotermi med den förväntade MR för en ektoterm av liknande storlek och är därför dimensionslös och oberoende av kroppsstorlek. (B) Den evolutionära vägen för minsta motstånd från ektotermi till endotermi inkluderar tröghetshomeotermi som ett övergångsstadium, följt av en ökning av ämnesomsättningen samtidigt med en minskning av storleken. (C) Hypotetisk sekvens av evolutionära övergångar i fågelstamlinjen, som kombinerar resultat från denna studie med fylogenetiska rekonstruktioner av epidermala strukturer och kapacitet för aktiv flygning (se huvudtexten). Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Det föreslagna scenariot förklarade varför teropoder systematiskt gynnade en dyr livsstil (högre energiomsättningshastighet) trots dess energikostnader, och ihållande miniatyrisering före fåglarnas ursprung. Vidare, ett tillstånd av tröghetshomeotermi utgjorde ett nödvändigt övergångstillstånd med låga metaboliska kostnader. Om stora förfäders theropoder var fysiologiskt engagerade i homeothermi, Rezende et al. förvänta dig sedan att de har avvikit från ektotermisk metabolisk allometri (samband mellan kroppsstorlek och form, anatomi, fysiologi och beteende) med miniatyrisering. Under evolutionär tid, det är rimligt att förvänta sig att härstamning ska utnyttja nyöppnade nischer och så småningom diversifiera sig. Resultaten av detta arbete är preliminära; de betonar helt enkelt att det sannolikt ligger mer bakom utvecklingen av endotermi hos arkosaurier, dinosaurier och dagens fåglar. Resultaten bildar en arbetshypotes som i framtida studier kan kombineras med tidigare analyser av utvecklingen av storlek och andra egenskaper i fågelstammar för att bilda väldefinierade tolkningar av tidssekvenser under viktiga evolutionära övergångar.
© 2020 Science X Network