Av Dianne Hermance | Uppdaterad 24 mars 2022
Andrew Brookes/Cultura/GettyImages
I eukaryota celler producerar mitos identiska dotterceller medan meios genererar genetiskt distinkta gameter. Båda processerna är beroende av exakt kromosomsegregation, en uppgift som utförs av spindelapparaten och dess fästen till kromosomerna.
Kinetochores och nonkinetochores mikrotubuli skiljer sig strukturellt och funktionellt, men de samarbetar för att säkerställa korrekt DNA-distribution under celldelning.
Mitos tillhandahåller nya celler för tillväxt, reparation och asexuell reproduktion. En ensamföräldercell delar sig i två genetiskt identiska döttrar genom att dela upp dess kärna och kromosomer.
Människor innehåller 23 kromosompar, vart och ett lagrat som två systerkromatider förenade vid centromeren. Att bibehålla kromosomtalet under delning är avgörande för genomisk stabilitet.
Celldelning styrs av interfas (G1, S, G2) och mitos, som börjar med profas. Under profas kondenserar kromatin till systerkromatider, kärnan löses upp och spindelapparaten bildas från mikrotubuli.
Prometafas följer, under vilken kärnhöljesfragmenten och spindelmikrotubuli fäster vid kinetokorer vid centromerer. I metafas kommer kromosomerna i linje med metafasplattan, och spindelmikrotubuli når mot motsatta poler.
Under anafas separeras systerkromatider och dras till motsatta poler av kinetochore-associerade mikrotubuli. Nonkinetochore mikrotubuli hjälper till med kromosomrörelser och spindelförlängning.
Telofas återställer kärnhöljen runt varje uppsättning kromosomer, och cytokines fullbordar processen genom att dela upp cytoplasman.
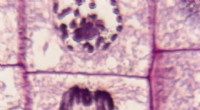
En kinetochore är ett stort multiproteinkomplex som samlas på centromeren av varje kromosom. Först beskrevs av Walther Flemming 1880, fungerar kinetokoren som fästställe för spindelmikrotubuli och en kontrollpunkt för korrekt kromosomjustering.
Trots artspecifika DNA-variationer är kinetokorarkitekturen mycket bevarad, vilket understryker dess grundläggande roll i mitos.
Kinetochores är strukturerade, proteinrika plattformar som överbryggar kromosomalt DNA och mikrotubuli, medan nonkinetochore mikrotubuli är dynamiska polymerer som underlättar spindelbildning och kromosomrörelse. De förra förankrar kromosomerna; den senare ger den mekaniska kraft som behövs för segregering.
Kinetochorer fungerar som mikroskopiska motorsammansättningar och omvandlar mikrotubulidynamik till riktade kromosomrörelser. De fungerar också som kvalitetskontrollpunkter; fel i vidhäftning utlöser AuroraB-kinasmedierad fosforylering, vilket stoppar utvecklingen till anafas tills den korrigeras.
Histonvarianten CENP-A kärnar centromeriskt kromatin och rekryterar CENP-C och andra inre kinetokorproteiner. Den yttre kinetokoren, som innehåller Ndc80-komplexet, kopplar direkt in mikrotubuli.
Under mitos genomgår kinetochore snabb montering och demontering, reglerad av fosforyleringshändelser. Ndc80-komplexet förankrar mikrotubuli, vilket gör att kinetochore kan "dansa" med de dynamiska plusändarna av mikrotubuli.
Motorproteiner som kinesin och dynein förbättrar kromosomrörelsen, medan mikrotubulusdepolymerisering vid kinetokorer genererar dragkrafter som segregerar kromatider.
Kinetochores övervakar fästets trohet. AuroraB-kinas upptäcker felaktiga mikrotubuli-kinetochore-bindningar och fosforylerar associerade proteiner, vilket leder till lösgörings- och återfästningsförsök. Komplex som Pcs1/Mde4 förhindrar ytterligare felkopplingar nära centromerer.
När fel kvarstår fördröjer spindelmonteringskontrollpunkten anafasstart, vilket ger tid för korrigering och säkerställer genomisk integritet.
Pågående forskning fortsätter att belysa den molekylära koreografin av kinetochores sammansättning och funktion. Framsteg inom kryoelektronmikroskopi och levande cellavbildning lovar djupare förståelse av kromosomsegregationsmekaniken i både mitos och meios.
Genom att reda ut dessa processer strävar forskare efter att upptäcka terapeutiska mål för sjukdomar som är förankrade i kromosomfelsegregering, såsom cancer och aneuploidsyndrom.