Konkurrens och samarbete är grundläggande krafter som styr den evolutionära och ekologiska dynamiken bland arter. Balansen mellan dessa krafter varierar mellan ekologiska sammanhang, med vissa miljöer som gynnar kooperativa beteenden som främjar ömsesidig nytta, medan andra belönar konkurrenskraftiga strategier som maximerar individuell kondition.
Bland mikrobiella samhällen ger kemikalier som utsöndras i miljön möjligheter till både samarbete och exploatering, vilket i vissa fall ger upphov till mikrobiella "fuskare". Dessa fuskare utnyttjar sina motsvarigheters samarbetsbeteende och drar nytta av de utsöndrade föreningarna utan att betala de metaboliska kostnaderna för produktionen.
I en ny artikel publicerad i Molecular Biology and Evolution , avslöjar forskare från University of Wisconsin-Madison och Vanderbilt University den evolutionära historien om utsöndrade järnupptagsmolekyler i jästsvampar, vilket kastar nytt ljus över den samarbets- och konkurrensdynamik som formar järnbegränsade mikrobiella samhällen.
De flesta organismer kräver järn för många biologiska processer men kan inte absorbera den vanligaste formen av järn i miljön. Järn är därför ofta en begränsad resurs i biologiska samhällen. För att övervinna denna brist har mikroorganismer utvecklat förmågan att ta bort järn från miljön med hjälp av sideroforer, molekyler med hög affinitet för den typ av järn som finns i miljön.
Sideroforer syntetiseras inuti cellen och utsöndras sedan i miljön, där de binder till järn; de järnbundna molekylerna måste sedan importeras tillbaka in i cellen innan järnet kan frigöras och användas i cellulär metabolism. Sideroforer som utsöndras i miljön kan utnyttjas av fuskare, som får en fitnessfördel genom att ta upp järnbundna sideroforer utan att investera energi i sin produktion.
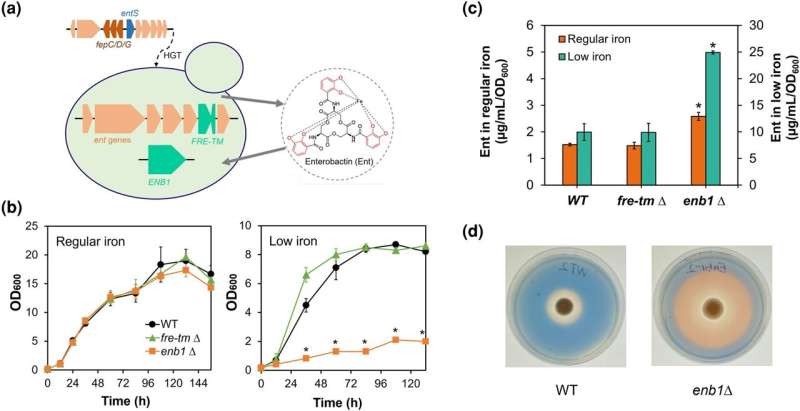
Medan de flesta jästsvampar inte kan producera sideroforer, fann ett forskarlag under ledning av Chris Hittinger att jästsvampar i Wickerhamiella/Starmerella (W/S) kladden kunde producera en siderofor som kallas enterobactin. Generna som krävdes för att syntetisera enterobaktin överfördes tydligen horisontellt från en gammal bakterie till förfadern till W/S-jästsvampar.
Spännande nog hade W/S-jästsvamparna inget uppenbart sätt att återimportera enterobaktinsideroforen när den väl var bunden till järn. "Vi hittade ingen bakteriegen som kodade för en enterobaktintransportör i deras genom", säger Liang Sun, huvudförfattare till den nya artikeln.
"Att utsöndra enterobactin utan att föra tillbaka det till cellen för järnupptag skulle inte vara ett smart drag för en jästcell, så vi var väldigt nyfikna på hur dessa jästsvampar skulle kunna utnyttja järnet bundet till enterobactin."
För att lösa detta pussel sökte teamet genomet av Starmerella bombicola efter en alternativ mekanism för siderofortransport. Genom riktade genstörningsexperiment och fylogenomiska analyser identifierade teamet en gen känd som ENB1 som avgörande för upptaget av enterobaktinbundet järn i St. bombicola. Överraskande nog är ENB1 en uråldrig svampgen som sannolikt går tillbaka hundratals miljoner år, före divergensen av svamphärkomsterna Basidiomycota och Ascomycota.
Ytterligare analyser avslöjade en komplex historia av ENB1 inom jäst. Forskarna föreslog att ENB1 överfördes horisontellt från en förfader till W/S-kladen till en uråldrig linje av Saccharomycetales, gruppen som inkluderar Saccharomyces cerevisiae, som används för att göra bröd, öl och vin. Denna överföring, tillsammans med efterföljande gendupliceringar och förluster, har format den ojämna fördelningen av enterobaktinanvändning som för närvarande observeras bland jästsvampar.
Dessa fynd har flera intressanta implikationer för historien om järnupptaget i jäst. Eftersom enterobaktinupptaget uppenbarligen går före förmågan att producera enterobactin i W/S-jäst, var förfäderna till denna kladd troligen fuskare som gynnades av produktionen av enterobaktin av andra mikrober i sin miljö.
Därefter förvärvade W/S-kladen enterobaktinbiosyntesgener från en bakterie inom ett ekologiskt sammanhang där det var mer fördelaktigt att vara producent än att vara en fuskare.
Baserat på vad som är känt om fördelningen av dessa jästsvampar, föreslår författarna till studien att detta inträffade i en insektstarm, där konkurrensen om järn mellan bakterier, jästsvampar och värden kan vara hård. Förmågan hos W/S-jäst att producera enterobactin och att importera det med Enb1-transportören kan ha gett en konditionsfördel i denna mycket konkurrenskraftiga, järnbegränsade miljö.
Däremot kan kvarhållandet av ENB1 hos fuskare som S. cerevisiae "förknippas med ekologiska nischer där bakterie- och svampsambor producerar enterobactin som svar på järnbrist", enligt studiens författare. "Omvänt kan förlusten av ENB1 ha inträffat i jästsvampar som lever i miljöer med relativt hög järntillgänglighet eller där enterobaktinproducenter saknas."
Även om dessa resultat är spännande, behövs ytterligare forskning för att helt avslöja mekanismerna genom vilka svamp- och bakterie enterobactingenerna integrerades i W/S-jästsvampar. Enligt Sun måste dessa gener vara hårt samreglerade, eftersom "obalanserad utsöndring och import av enterobaktin kan hindra järnupptaget och därefter leda till tillväxtdefekter i jästsvamparna."
Tyvärr noterar Sun att de metaboliska och reglerande nätverken för dessa jästsvampar inte är väl förstådda, vilket kan göra framtida studier utmanande, "Att studera regleringen av denna speciella väg kan därför kräva ytterligare ansträngning för att fylla i några av dessa luckor."
Trots dessa hinder erbjuder detta system en unik modell för vidare forskning om den evolutionära dynamiken hos siderofortransportörer i jästsvampar och deras roll för att främja samarbete och fusk inom mikrobiella samhällen.
Mer information: Liang Sun et al, Funktionell och evolutionär integration av en svampgen med en bakteriell operon, Molecular Biology and Evolution (2024). DOI:10.1093/molbev/msae045
Journalinformation: Molekylärbiologi och evolution
Tillhandahålls av Society for Molecular Biology and Evolution