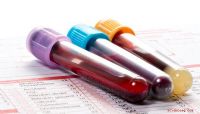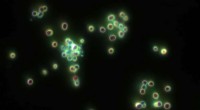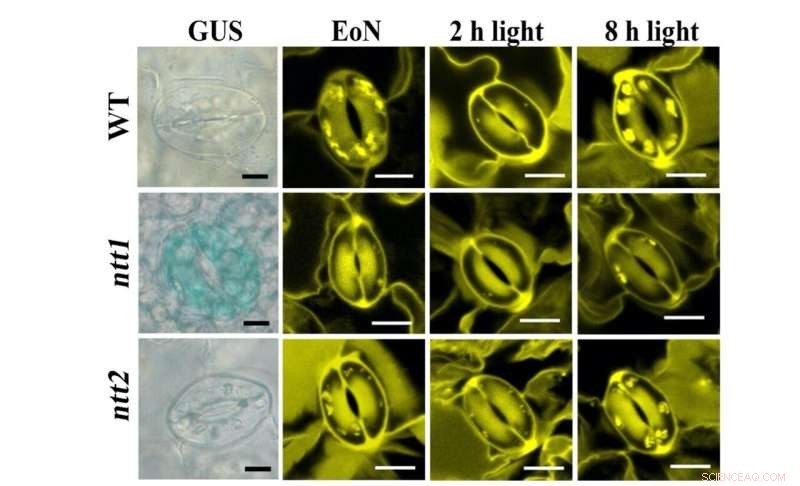
Figur 1:Färgning av NTT-transportörer (blå, vänster panel) och stärkelsegranulat (gul, höger panel) i GCs. Kredit:University of Hong Kong
Huruvida Guard Cells (GC) utför fotosyntes har diskuterats i decennier. Tidigare studier antydde att skyddscellkloroplaster (GCC) inte kan fixera CO2 men senare studier hävdade något annat. Tills nyligen har det varit kontroversiellt om GCC och/eller GC-fotosyntes spelar en direkt roll i stomatala rörelser. Dr. Boon Leong LIM, docent vid School of Biological Sciences vid University of Hong Kong (HKU), upptäckte i samarbete med Dr. Diana SANTELIA från ETH Zürich GCs verkliga bränslekälla och red ut mysteriet. Resultaten publicerades nyligen i den prestigefyllda tidskriften Nature Communications .
På morgonen utlöser solljus stomata, som är små porer på växtens blad, att öppna sig. Detta låter CO2 in och O2 för att öka fotosyntesen. Öppnandet av stomata förbrukar en stor mängd adenosintrifosfat (ATP), den cellulära energivalutan, men källorna till ATP för stomatöppning förblev oklara. Vissa studier antydde att GCC utför fotosyntes och exporterar ATP till cytosolen för att aktivera stomatöppningen. I mesofyllkloroplaster genereras ATP och NADPH (nikotinamid-adenindinukleotidfosfat) från fotosystem, som används som bränsle för att fixera CO2 .
Genom att använda i planta fluorescensproteinsensorer kunde teamet till Dr. Boon Leong Lim vid HKU visualisera realtidsproduktion av ATP och NADPH i mesofyllcellkloroplasterna (MCC) i en modellväxt, Arabidopsis thaliana. "Vi kunde dock inte upptäcka någon ATP- eller NADPH-produktion i GCC under belysning. Förbryllade över denna oväntade observation kontaktade vi en expert på vaktcellsmetabolism, Dr. Diana Santelia från ETH Zürich, för ett samarbete," sa Dr. Lim. Under det senaste decenniet har Santelia-labbet gett djupa och viktiga insikter om stärkelse- och sockermetabolism i skyddscellerna (GC) som omger stomatala porer på bladytan.
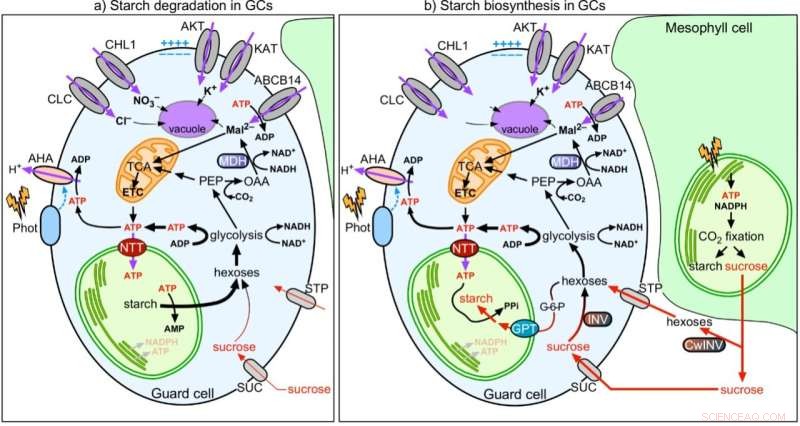
Figur 2:Modell av koordinationen av stomatal funktion med stärkelse- och malatmetabolism i skyddsceller. Kredit:University of Hong Kong
I gemensamma ansträngningar visar teamet att till skillnad från mesofyllceller (MC) är GC-fotosyntesen dåligt aktiv. Socker som syntetiseras och levereras av MC importeras till GC och konsumeras av mitokondrier för att generera ATP för stomatal öppning. Till skillnad från MCC tar GCC upp cytosolisk ATP via nukleotidtransportörerna (NTT) på kloroplastmembranet för att aktivera stärkelsesyntesen på dagtid. I gryningen, medan MC börjar syntetisera stärkelse och exportera sackaros, bryter GC ner stärkelse till sockerarter för att tillföra energi och öka turgortrycket för stomatal öppning. Följaktligen är funktionen hos GCC att fungera som ett lager av stärkelse viktig för stomatal öppning. Medan MC fixar CO2 i kloroplaster via cykeln Calvin–Benson–Bassham (CBB), CO2 fixering i cytosolen är den huvudsakliga vägen för CO2 assimilering i GC, där nedströmsprodukten malat, är också ett viktigt löst ämne för att öka turgortrycket för stomatal öppning. Sammanfattningsvis beter sig GC mer som ett handfat (ta emot sockerarter) än en källa (tillhandahåller sockerarter) vävnad. Deras funktion är nära korrelerad med den för MC:er för att effektivt koordinera CO2 upptag via stomata och CO2 fixering i MCs.
"Jag blev väldigt upprymd när Dr. Lim kontaktade mig och bad om att få samarbeta i detta projekt," sa Dr. Diana Santelia. "Vi har försökt klargöra dessa grundläggande frågor med hjälp av molekylärgenetiska metoder. Att kombinera vår respektive expertis har varit en vinnande strategi", fortsatte hon. Dr. Sheyli LIM, artikelns första författare och tidigare Ph.D. elev i Lims grupp anmärkte "De in planta fluorescensproteinsensorer vi utvecklade är kraftfulla verktyg för att visualisera dynamiska förändringar av koncentrationerna av energimolekyler i individuella växtceller och organeller, vilket gör att vi kan lösa några nyckelfrågor inom växtbioenergetik. Jag är glad att publicera våra upptäckter i Nature Communications använder den här nya tekniken."